 |
|
| Background Pectins are a family of complex polysaccharides that contain 1,4-linked α-D-galactosyluronic acid residues. Pectins account for ~30% of the primary walls of dicotyledenous and non-graminaceous monocotyledenous plants and for between 5 and 10% of the walls of grasses. Pectins are also likely to be present in the walls of ferns, lycopods, and bryophytes. The presence of pectin in algal walls needs to be substantiated. There is no evidence that bacteria or fungi produce pectin. Three pectic polysaccharides have been isolated from plant primary cell walls and structurally characterized. These are:
Specific pages are devoted to Rhamnogalacturonan I and Rhamnogalaturonan II |
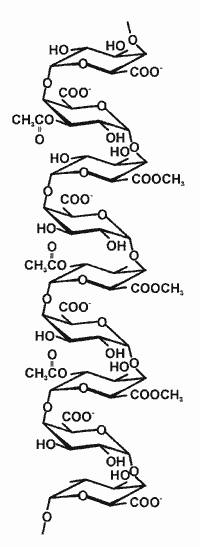
| Homogalacturonan Homogalacturonan (HG) is a linear chain of 1,4-linked α-D-galactosyluronic acid residues in which some of the carboxyl groups are methyl esterified (see the figure on the right). HG with a high degree of methyl esterification is referred to as "pectin" whereas HG with a low degree of methyl esterification is "pectic acid". HGs may, depending on the plant source also be partially O-acetylated. HG may account for up to 60% of the pectin in a primary wall. Many of the properties and biological functions of HG are believed to be determined by ionic interactions. Increases in the degree of methyl esterification of HG have been implicated in increased cell separation. Enzymes that fragment HG (endo and exo polygalacturonases and lyases) are secreted by many plant pathogens. The oligogalacturonide fragments generated have been shown to induce several plant defence responses (see Plant Pathogens). The sequencing of the Arabidopsis genome has revealed the existence of at least 52 genes encoding putative polygalacturonases. However, there is little know about the function or specificities of these enzymes. Studies with transgenic plants that over-express a specific endopolygalacturonase (EPG) or have had the levels of endogenous EPG reduced have begun to provide information about the function of HG. For example, neither the supression or over-expression of EPG in tomato fruits affected the fruit ripening process but did alter tissue texture and pectin solubility. In contrast transgenic apples containing additional copies of a fruit-specific EPG exhibited premature leaf shedding, reduced cell adhesion, brittle leaves, and had malformed stomata. It is likely that HG together with other pectic polysaccharide have numerous roles in plant growth and development HG biosynthesis is a major area of research at the CCRC (see Biosynthesis) |
 |
| Unusual ester cross links in pectin The existence of esters that form between the carboxyl group of a 4-linked GalpA residue and a hydroxyl group of another glycosyl residue has been hypothesized. The presence of non-methyl galacturonosyl esters has been implied from chemical analysis and FT-IR spectroscopy. The trans-esterification of pectin has been reported to be catalyzed by pectin methyl esterases. However, no fragments of HG containining unusual esters have been isolated and structurally characterized. |
| Current research 1.The use of chemical and enzymic methods to isolate and characterize non-methyl esters in pectin. |
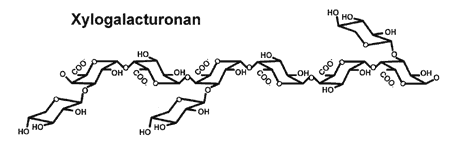
| Xylogalacturonan Xylogalacturonans have β-D-Xylp residues attached to C-3 of the galacturonan backbone (see right figure). Xyloglacturonans are only present in reproductive tissues including soybean seed, apple fruit, and pine pollen. Little is known about the function of this polysaccharide, although it has been shown that the presence of Xylp residues limits the ability of fungal EPGs to fragment the backbone |
 |
|
Apiogalacturonan |
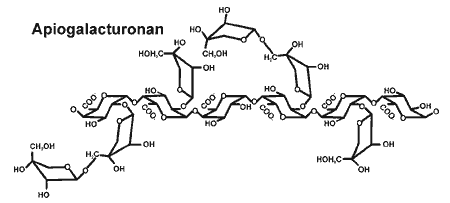 |
| Current research 1. The biosynthesis of apiose and xylose - further details are available in the biosynthesis page. |
| Selected References ONeill et al (1990) The pectic polysaccharides of primary cell walls. In Methods in Plant Biochemistry (Dey and Harborne eds) Vol 2, Academic Press, London, pp 415-441. Schols et al (1995) A xyloglacturonan subunit present in the modified hairy regions of apple pectin. Carbohydr Res., 279, 265-279 Ridley et al (2001) Pectins: structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry, 57, 929-967 Golovchenko et al (2001) Structural studies of the pectic polysaccharide from duckweed Lemna minor L. Phytochemistry, 60, 89-97. Atkinson et al (2002) Over expression of polygalacturonase in transgenic apple trees leads to a range of novel phenotpyes involving changes in cell adhesion. Plant Physiol., 129, 122-133. |


